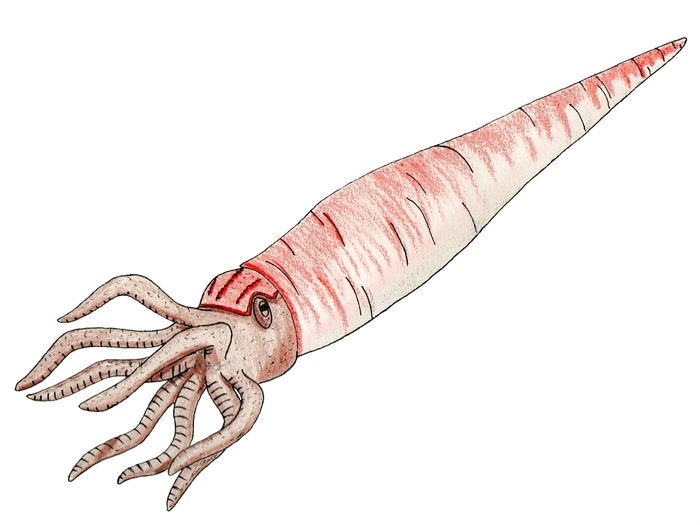
Раковина камероцераса достигала длины 9—10 м, то есть, вместе с щупальцами этот моллюск должен был иметь длину 11 метров (позднее эти данные пересмотрены в меньшую сторону). Этот моллюск был одним из крупнейших животных, живших в эпоху палеозоя. Судя по его огромным размерам, он был высшим хищником, обитавшим на глубоководье и, вероятно, питавшимся ракоскорпионами, такими как мегалограпт (Megalograptus welchi), большими трилобитами, и небольшими головоногими.
Камероцерас стал мусорным таксоном, к этому роду могут отнести любых крупных ортоконов, таких как эндоцерас (Endoceras), вагиноцерас (Vaginoceras), менискоцерас (Meniscoceras). Хотя Cameroceras trentonense был впервые описан Конрадом в 1842 году, с тех пор термин использовался в разных значениях. Cameroceras и Endoceras используется даже для описания разных стадий развития одного вида.
Амфибия достигала в длину от 35 см до, вероятно, 1 м. Крупная округлая голова, большие глаза, мощные ноги, относительно короткие тело и хвост. Борозд боковой линии нет. Очень большая «ушная вырезка» — многие авторы предполагают наличие барабанной перепонки и хорошо развитый слух (и даже наличие вокальных данных, как у лягушек). Однако, стремя, по-видимому, массивное. Тело, в отличие от современных земноводных, покрыто мелкими чешуйками. Наземный хищник.
Скелет Dendrerpeton acadianum
По данным сайта Paleobiology Database на ноябрь 2023 года в род включают 4 вымерших вида:
Размеры тела достигали 2 метров. У рыб была большая плоская голова с мощным ртом и маленькими глазами. Предположительный образ жизни: придонные хищники, подстерегавшие жертву в засаде.
В локации Лоде (Латвия) было найдено захоронение среднего девона, где представлены полные скелеты Laccognathus panderi вместе с пластинокожими Asterolepis ornata и лопастепёрыми рыбами Panderichthys. Все эти животные погибли в момент максимального пересыхания речной дельты, где они обитали.
† Laccognathus embryi Daeschler & Down, 2011 — в длину достигал 1,8 метров. Ископаемые остатки были обнаружены в алевролитовых пойменных отложениях на острове Элсмир в Нунавуте, канадская Арктика.
† Laccognathus grossi Vorobyeva, 2006
† Laccognathus panderi Gross, 1941
J. Mondéjar, Fernández, M. Friedman, and S. Giles. 2020. Redescription of the cranial skeleton of the Early Devonian (Emsian) sarcopterygian Durialepis edentatus Otto (Dipnomorpha, Porolepiformes).
Скелет тиктаалика в Филдовском музее естественной истории
Реконструкция
Находка была сделана на канадской территории Нунавут, где обитают эскимосские племена. В 2006 году Нил Шубин послал Совету старейшин территории просьбу выбрать для находки имя. Совет старейшин предложил два варианта: Siksagiaq и Tiktaalik. Из соображений удобопроизносимости был выбран второй вариант, который на языке инуктитут означает «налим».
Тиктаалик является переходным звеном между рыбами и наземными позвоночными. В его строении сочетаются черты тех и других.
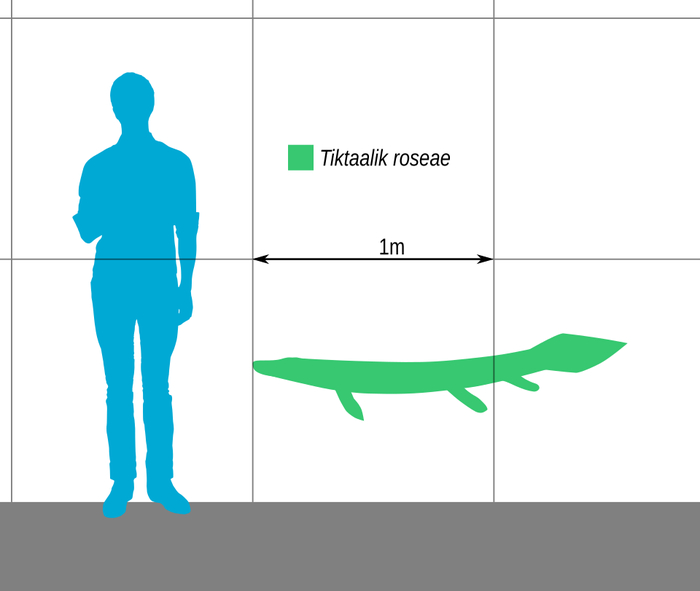
Сравнительный размер Тиктаалика
Признаки, свойственные рыбам: жабры и чешуя.
Признаки, характерные для переходных форм от рыб к четвероногим: строение костей и суставов конечностей, в том числе функциональный лучезапястный сустав, свойственный четвероногим, при наличии плавников, а также строение уха.
Признаки, традиционно приписываемые четвероногим: рёбра, по строению схожие с рёбрами четвероногих; подвижный шейный отдел; лёгкие.
Согласно палеонтологическим данным, находки самых ранних полностью сформировавшихся амфибий относятся к отложениям возрастом 365 млн лет. В отложениях возрастом 385 млн лет до сих пор не было обнаружено никаких следов животных, которых можно классифицировать как переходные формы между рыбами и земноводными. Таким образом, наиболее вероятно нахождение переходных форм в отложениях возрастом около 375 млн лет. На североамериканском континенте известно три места, где отложения этого периода, относящиеся к дельтам крупных рек, выходят на поверхность. Они находятся в штате Пенсильвания (США), Восточной Гренландии и на арктических островах Канады.
Район Восточной Гренландии к началу 2000-х годов был хорошо изучен. Именно здесь Дженни Клек из Кембриджского университета нашла самое древнее известное земноводное. В штате Пенсильвания на месте взрывных работ при строительстве дороги Нил Шубин (один из первооткрывателей тиктаалика) нашёл лопатку, относящуюся к древнему земноводному Hynerpeton возрастом 360 млн лет. Территория же канадских арктических островов была совершенно не исследована палеонтологами. Именно из этих соображений Нил Шубин в 1999 году предпринял экспедицию на остров Мелвилл на крайнем западе канадской Арктики[4].
Поверхность острова Мелвилл представляла собой отложения, сформировавшиеся на дне древнего океана. Здесь было найдено множество останков глубоководных рыб, однако найти здесь переходные формы оказалось невозможным. Нужны были отложения древних озёр и речных мелководий, поэтому в 2000 году экспедиция переместилась восточнее — на остров Элсмир, который 375 млн лет назад представлял собой побережье древнего континента Лаврентия и располагался в районе экватора. В конце полевого сезона 2000 года экспедицию ждала удача — был найден слой с большим количеством останков рыб. Но только в 2004 году исследователи обнаружили ископаемые остатки рыбы с плоской уширенной головой, передние конечности которой были сформированы сходно с тетраподами — к лопатке крепилось подобие плечевой кости, затем парные кости, соответствующие лучевой и локтевой, затем множество костей, сходных с запястьем и пальцами. Возраст находки составил 375 млн лет.
*Тут должна быть мультипликационная реконструкция локомоции но пикабу не может загружать гифки(*
Тело уплощённое, голова широкая, несколько сужена кпереди. Вентральный щит панциря очень небольшой, сдвинут под переднюю часть головы. Грудной панцирь узкий, постбранхиальные пластинки (соединяющие спинной и брюшной щиты) в виде бивнеобразных отростков, загнутых вперёд. Сам грудной панцирь несколько скошен назад, в результате чего морда едва выступает за пределы брюшного щита. Глаза очень маленькие, у переднего края головы, вынесены наверх. Швы головного щита сложные, карманообразные. Хорошо развиты борозды боковой линии. Челюсти плохо известны, вероятно, слабые. Наружная структура панциря бугорчатая. На внутренней поверхности непарной спинной пластины грудного панциря развит мощный направленный назад каринальный отросток, к которому крепились мышцы спины. Размеры огромные — поперечник панциря более 1 метра, общая длина до 6 метров.
Придонные пресноводные рыбы. Образ жизни не вполне понятен. Возможно, питались беспозвоночными.
Род предложен Х. Асмуссом в 1856 году на основании остатков из среднего девона (живета) Эстонии — пещеры Арукюла близ города Тарту. Синонимы — Chelonichthys, Heterostius, Ichthyosauroides (некоторые остатки ошибочно относили к рептилиям, в частности, С. С. Куторга). Луи Агассис причислял типовые остатки к роду Asterolepis. Тип рода — Heterosteus asmussi. Возможный синоним типового вида — H. ingens. Несколько видов, остатки известны из Ленинградской области, Рейнской области Германии, Шпицбергена и, возможно, Гренландии. Из среднего девона Шпицбергена известен родственный, но несколько более мелкий Herasmius, а из среднего девона Китая (Юньнань) — Yinostius.
Основы палеонтологии: Бесчелюстные и рыбы / под ред. Д. В. Обручева. — М., 1964. — С. 154—155.
Черепанов Г. О., Иванов А. О. Палеозоология позвоночных. — М., «Академия», 2007. — С. 91.
Janvier Ph. Early Vertebrates. — Oxford Science Publications. — 2003. — P.160—161.
Арандаспиды (лат.Arandaspida) — небольшая группа вымерших панцирных бесчелюстных из класса парноноздрёвых (Pteraspidomorpha), которой присваивают ранг от отряда до подкласса. Название образовано от названия австралийского народааранда и др.-греч. ἀσπίς — «щит» (обычное окончание для названий вымерших бесчелюстных).
По внешнему виду арандаспиды были рыбоподобными животными, но имели только один плавник — хвостовой. Достигали длины 35 см. Были покрыты тонким панцирем. Примечательны парностью отверстия для теменного глаза (хотя эти отверстия имеют и другую интерпретацию).
В составе этой группы различают от 2 до 8 видов. Арандаспид сближают с разнощитковыми, а иногда и включают в их состав.
Чаще всего к арандаспидам относят 4 рода, 2 из которых — предположительно, поскольку они известны только по мелким фрагментам. В каждом из этих родов описан только один вид:
Sacabambaspis janvieri Gagnier et al., 1986, известный из Боливии, Аргентины, Австралии и Омана. Назван в честь типового местонахождения — боливийского города Сакабамба. Жил в конце нижнего ордовика (лланвирнский и карадокский век, около 470 млн лет назад). Известен по останкам намного лучшей сохранности, чем остальные арандаспиды, в том числе по практически целым экзоскелетам.
Andinaspis suarezorum Gagnier, 1991 из ордовика или девона Боливии (недалеко от местонахождения Sacabambaspis). Известен по единственному обломку; принадлежность к арандаспидам не доказана.
Arandaspis prionotolepis Ritchie & Gilbert-Tomlinson, 1977 из центральной Австралии (Алис-Спрингс). Назван в честь живущей в этих местах этнической группы. Жил в начале верхнего ордовика (карадокский век, около 450 млн лет назад). Известен по отпечаткам передней части экзоскелета в мелкозернистом песчанике;
Porophoraspis crenulata Ritchie & Gilbert-Tomlinson, 1977 оттуда же. Кроме того, подобные ему окаменелости известны из раннеордовикских пород (аренигский ярус, около 480 млн лет назад), и это самые древние находки арандаспид). Получил название за большие поры на бугорках панциря. Известен только по мелким фрагментам; принадлежность к арандаспидам не доказана.
Кроме того, в родах Sacabambaspis, Arandaspis и Porophoraspis, вероятно, есть как минимум по одному неописанному виду. Не исключено, что к арандаспидам относится и ещё один боливийский вид — Pirchanchaspis rinconensis Erdtmann et al., 2000, но его систематическое положение очень неясное.
Почти целая окаменелость
Арандаспиды имели вытянутое каплевидное тело длиной до 35 см, покрытое хорошо развитым экзоскелетом. Брюшная сторона была более выпуклой, спинная — более плоской. Единственный плавник — хвостовой. Он хорошо изучен только у Sacabambaspis, у которого имел длинную, но узкую центральную лопасть (куда заходила хорда), более короткую спинную и ещё меньшую брюшную.
Глаза находились на самом конце головы. В них были окостенения — склеротические кольца. Между глазами расположены ноздри, а позади — пара отверстий, которые обычно интерпретируются как пинеальные.
Рот арандаспид был на нижней стороне головы. У них было до 10 пар жаберных мешков, которые открывались наружу отдельными отверстиями, а по другим данным — в общие проходы, открывавшиеся одним отверстием с каждой стороны.
Реконструкция Сакабамбасписа
Эндоскелета у арандаспид не было или почти не было, зато был хорошо развит экзоскелет. Переднюю часть тела покрывали две большие костные пластины (спинная и брюшная), а заднюю — сильно вытянутые вертикальные чешуйки. Длина передних пластин составляет около половины длины животного, а толщина очень маленькая (у Arandaspis < 0,1 мм). Они цельные (без следов слияния отдельных элементов) и не имеют признаков роста. Это означает, что они появлялись уже у взрослого животного.
Между спинной и брюшной пластиной с каждой стороны был ряд из 15—20 небольших многоугольных пластинок, между которыми находились жаберные отверстия. Кроме того, много рядов мелких костных пластинок было с нижней стороны рта. На спинном и брюшном щитке, а также на мелких чешуйках видны маленькие, но хорошо развитые каналы боковой линии.
Все элементы экзоскелета образованы аспидином — бесклеточной костью. В них различаются 3 слоя: нижний (пластинчатый), средний (сотоподобный или сетчатый) и верхний (бугорчатый).
Снаружи панцирь был орнаментирован бугорками. Для родов, известных только по обломкам (Andinaspis и Porophoraspis) их форма — главный отличительный признак. У Arandaspis они вытянутые, сужаются к концам и имеют продольный гребень, а у Sacabambaspis, Andinaspis и Porophoraspis в разной мере напоминают дубовые листья, причём у последнего пронизаны большими порами.
Формой и орнаментацией панциря арандаспиды напоминают разнощитковых и астраспид, а чешуйками хвостовой части — анаспид.
Позади глазниц арандаспид, в передней части спинного щита, расположена пара небольших отверстий. Они есть и у Arandaspis, и у Sacabambaspis. Обычно их интерпретируют как отверстия для органов пинеального комплекса — светочувствительной структуры, из которой у многих древних и некоторых современных позвоночных развивается теменной глаз. Таким образом, у арандаспид таких глаз было два: одно отверстие вмещало пинеальный орган, а другое — парапинеальный. Это очень редкая ситуация среди позвоночных (кроме арандаспид, парное или сдвоенное пинеальное отверстие известно только у некоторых ископаемых рыб — ряда плакодерм, палеонисцид, поролепообразных и ранних двоякодышащих). Кроме того, глазоподобность обоих органов пинеального комплекса сохранилась и у большинства миног (хотя пинеального отверстия у них нет).
По другой интерпретации, эти отверстия у арандаспид представляют собой выходы эндолимфатических протоков, которые есть на панцире и некоторых других остракодерм. В пользу этого говорит то, что они расположены дальше назад, чем обычно для пинеального отверстия.
Арандаспиды жили в мелком море. Поскольку они не имели стабилизирующих плавников, их движение, вероятно, было неуклюжим и напоминало движение головастика. Тем не менее они были самыми прогрессивными известными позвоночными своего времени.
Как и для других остракодерм, для арандаспид предполагается придонный образ жизни и питание детритом и микроорганизмами. На это указывает положение рта снизу головы и (как и у других остракодерм) отсутствие челюстей.
Арандаспиды — древнейшие позвоночные, которые известны по довольно полным скелетам (от более древних находили только мелкие обломки панциря или отпечатки бесскелетного тела). Самые древние фрагменты, вероятно, принадлежащие арандаспидам, имеют возраст около 480 млн лет (начало ордовика), а самые молодые — 440 млн лет (конец ордовика). Причина их вымирания неизвестна; возможно, они исчезли из-за оледенения.
В отличие от других ранних остракодерм (живших в основном в водах Лаврентии), арандаспиды известны только из Гондваны (а именно из прибрежных отложений окружавших её морей). Окаменелости этих животных встречаются в Южной Америке (Боливия и Аргентина), Австралии (центр континента) и на Аравийском полуострове (Оман). Эти места были далеки друг от друга и в ордовике, хотя все находились на краю Гондваны.
Гондвана. Находки арандаспид известны с восточного, западного и северо-восточного края.
Окаменелости арандаспид встречаются редко, но к этой группе относятся почти все ордовикские остракодермы Гондваны. В водах этого континента остракодермы были редки и в последующие времена. Кроме арандаспид, оттуда известны только телодонты (очень широко распространённая группа, возникшая, видимо, в водах Лаврентии и к концу ордовика проникшая в моря Гондваны) и питуриаспиды (маленькая эндемичная для Австралии группа из раннего — среднего девона).
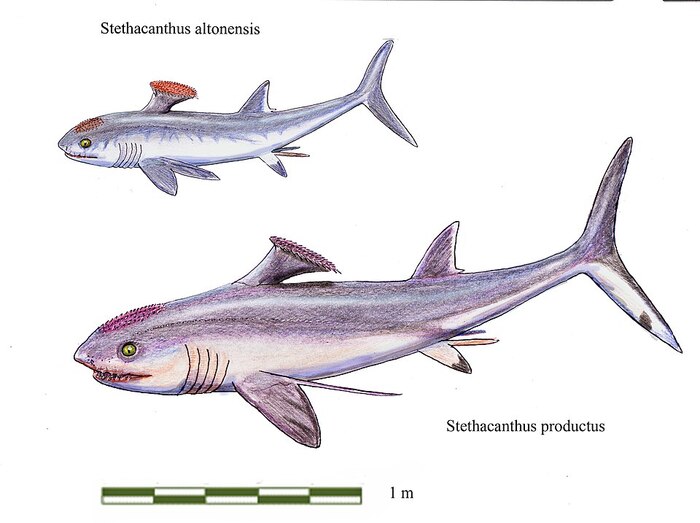
Достигали длины 0,7—2 м. Из-за диковинной формы спинного плавника стетакантов в шутку называют «гладильная доска». Только самцы обладали причудливым спинным плавником, а у самок плавник был обычным, как у современных акул. Назначение этого странного спинного плавника, как и коротких шипов на голове и передних плавниках, неизвестно. Возможно, он играл роль в привлечении самок.
По данным сайта Paleobiology Database, на август 2021 года в род включают 12 вымерших видов:
Stethacanthus altonensis (St. John & Worthen, 1875) [syn. Physonemus altonensis St. John & Worthen, 1875]
Stethacanthus compressus Newberry, 1897
Stethacanthus concavus Ginter, 2018
Stethacanthus depressus (St. John & Worthen, 1875) [syn. Physonemus depressus St. John & Worthen, 1875]